La lipidation de LGG-1/GABARAP n’est pas nécessaire pour l’autophagie et le développement chez Caenorhabditis elegans
Lien vers l'article originalAuteurs
Romane Leboutet, Céline Largeau, Leonie Müller, Magali Prigent, Grégoire Quinet, Manuel S. Rodriguez, Marie-Hélène Cuif, Thorsten Hoppe, Emmanuel Culetto, Christophe Lefebvre and Renaud Legouis
Année de publication
2023
Journal
eLife
Affiliation
Université Paris-Saclay, CEA, CNRS, Institute for Integrative Biology of the Cell (I2BC), 91198, Gif-sur-Yvette, France. INSERM U1280, 91198, Gif-sur-Yvette, France
Résumé de l'article
Les protéines de type ubiquitine Atg8/LC3/GABARAP sont nécessaires à de multiples étapes de l'autophagie, telles que l'initiation, la reconnaissance et la séquestration du cargo, la fermeture de la vésicule et sa dégradation. La plupart des fonctions de LC3/GABARAP sont considérées comme dépendantes de leurs modifications post-traductionnelles et de leur association avec la membrane de l'autophagosome via sa conjugaison à un lipide, la phosphatidyl-éthanolamine. Contrairement aux mammifères, C. elegans possède des homologues uniques des familles LC3 et GABARAP, nommés LGG-2 et LGG-1. Par mutagenèse dirigée, nous avons inhibé la conjugaison de LGG-1 à la membrane de l'autophagosome et généré des mutants qui n'expriment que les formes cytosoliques, soit le précurseur, soit la protéine clivée. LGG-1 est un gène essentiel pour l'autophagie et le développement chez C. elegans, mais nous avons découvert que ses fonctions pouvaient être pleinement réalisées indépendamment de sa localisation à la membrane. Cette étude révèle un rôle essentiel de la forme clivée de LGG-1 dans l'autophagie mais aussi dans une fonction embryonnaire indépendante de l'autophagie. Nos données remettent en question l'utilisation de la GABARAP/LC3 lipidée comme marqueur principal du flux autophagique et soulignent la grande plasticité de l'autophagie.
Image d'illustration
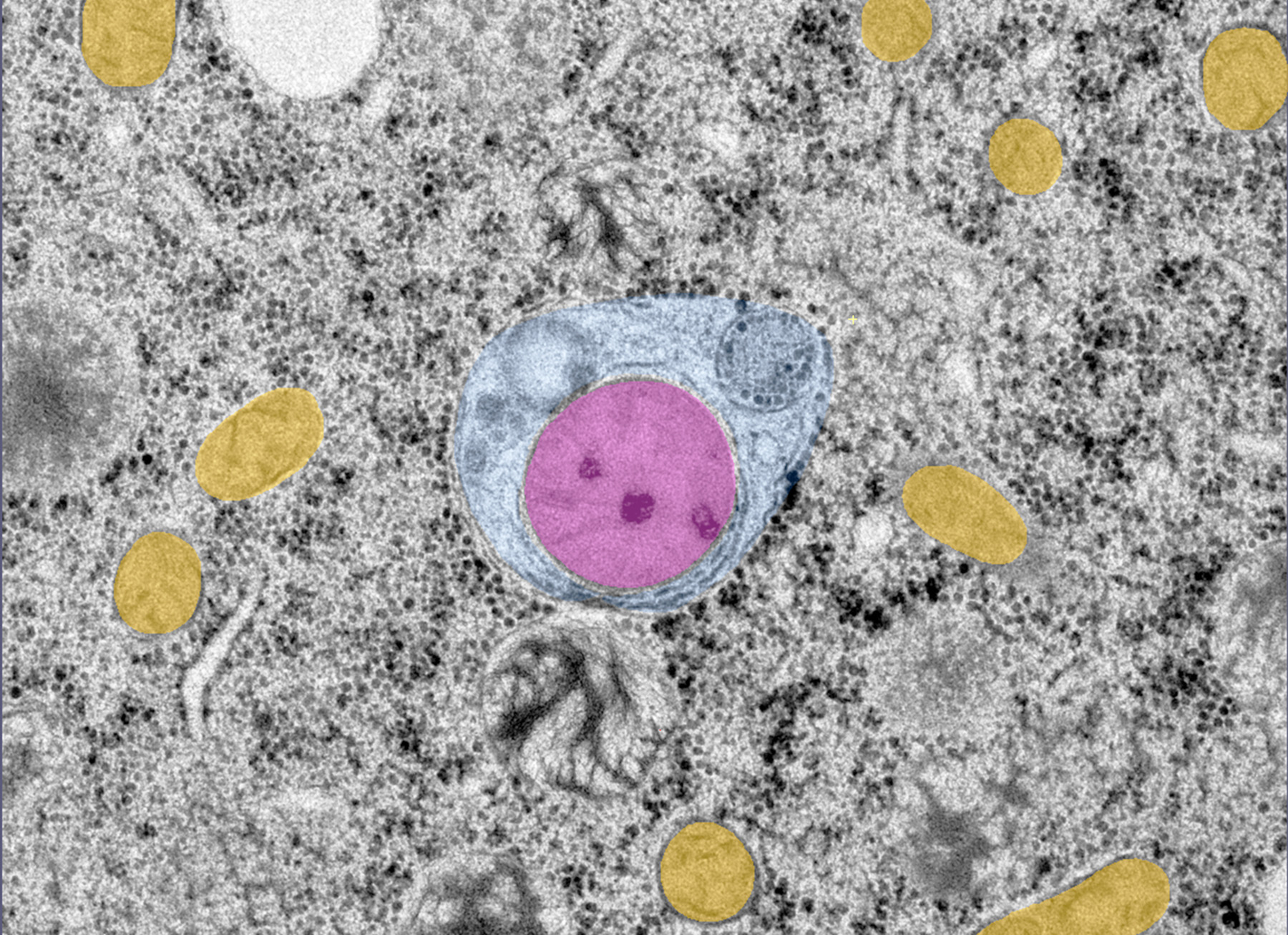
Microscopie électronique d'un embryon de C. elegans montrant un autophagosome (bleu) contenant une mitochondrie paternelle (rose) et entouré de mitochondries maternelles (jaune).